
Движение воды в радиальном направлении из почвенного раствора в проводящие элементы ксилемы осуществляется как непосредственно через клетки, так и по апопласту тканей корня (рис. 9).
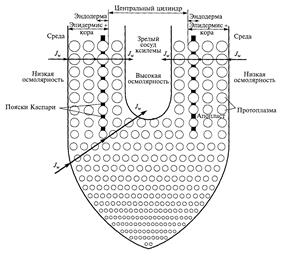
Рис. 9 Транспортная модель корня
(по E. Steudle and C.A. Peterson, 1998)
Апопласт представлен единой системой клеточных стенок и межклеточных полостей (межклетников). Транспорт воды через клетки получил название движение воды от клетки к клетке («cell to cell»). В данном случае вода может идти двумя путями: может пересекать мембраны и идти по непрерывному цитоплазматическому пути через соединяющие соседние клетки плазмодесмы. Единая система, объединяющая с помощью плазмодесм протопласты растительных клеток, называется симпластом. Таким образом, путь движения воды от клетки к клетке подразделяется на трансмембранный и симпластический. В трансмембранном движении воды важную роль играют аквапорины, образующие водные каналы. В итоге, движение воды от клеток ризодермы к сосудам ксилемы корня может осуществляться по трем параллельным направлениям: 1) через клеточные мембраны (трансмембранный путь); 2) по цитоплазме и плазмодесмам (симпластный путь); 3) по клеточным стенкам и межклетникам (апопластный путь). Эти три пути движения воды неавтономны: вода может перемещаться по комбинированным траекториям. Например, пройти некоторый путь по апопласту, затем пересечь плазматическую мембрану и далее передвигаться от клетки к клетке по симпласту.
В настоящее время отсутствуют экспериментальные подходы, которые позволили бы количественно оценить трансмембранный и симпластный потоки воды в корне. Однако имеющиеся данные позволяют предположить, что трансмембранные потоки значительно превосходят симпластные. Из-за отсутствия возможности разделить эти два потока, как правило, пользуются упрощенной транспортной моделью, в которой рассматривают два параллельных пути – путь от клетки к клетке и апопластный путь. Оба потока существуют в растении одновременно. Вклад того и/или иного пути в суммарный поток воды определяется многими факторами: видом растения, его возрастом, условиями выращивания и др.
При апопластном транспорте вода ни разу не пересекает мембраны, а сам апопласт не обладает свойством полупроницаемости. Это означает, что осмотические явления не вовлечены в формирование движущих сил апопластного транспорта воды. В этой связи ток воды через апопласт имеет гидравлическое происхождение, так как движущей силой для него служит преимущественно градиент гидростатического давления (∆Р), и ток воды по апопласту называется гидравлическим. Основным процессом, приводящим к формированию ∆Р, при апопластном (гидравлическом) транспорте является транспирация. При отсутствии водного дефицита в дневное время, когда устьица открыты и интенсивность транспирации высока, поток воды через апопласт в значительной степени превышает поток через клетки. При низкой интенсивности транспирации преобладающим становится движение воды от клетки к клетке.
При движении воды от клетки к клетке, когда вода на своем пути пересекает мембраны, явление осмоса играет определяющую роль. Ток воды в этом случае называется осмотическим, хотя градиент водного потенциала для него включает не только осмотическую, но и гидравлическую составляющую. Градиент водного потенциала между двумя клетками равен:
где ∆π, ∆Р – разности тургорного и осмотического давления между двумя клетками;
δ – коэффициент отражения мембран (он стремится к единице, поэтому ∆Ψ = ∆Р — ∆π).
Следует обратить внимание на то, что ∆Р при апопластном токе – это градиент гидростатического давления, создаваемого натяжением жидкости в сосудах ксилемы, а ∆Р при транспорте воды от клетки к клетке – это градиент тургорного давления в клетках корня.
На пути радиального транспорта воды в корне по апопласту расположена эндодерма с поясками Каспари. Основным химическим компонентом поясков Каспари является не суберин, как полагали ранее, а лигнин. Лигнин – это гидрофильное соединение, поэтому он не может препятствовать апопластному пути движения воды. До недавнего времени считали, что движение воды по апопласту в радиальном направлении корня прерывается на уровне эндодермы. Поэтому, чтобы продолжить свое движение в радиальном направлении корня, вода должна перейти в симпласт. Некоторое количество суберина в поясках Каспари может уменьшить гидравлическую проводимость. Таким образом, апопласт эндодермы не оказывает существенного влияния на водный ток, однако является препятствием для движения ионов. Так, на молодых корнях кукурузы было показано, что повреждение эндодермы приводит к выходу ксилемного раствора через повреждение из компартмента с высоким осмотическим давлением (центральный цилиндр) в компартмент с низким давлением (внешняя среда). Эти результаты подтвердили давно предполагавшуюся роль эндодермы как барьера для обратной диффузии ионов из центрального цилиндра в апопласт тканей коры.
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Да какие ж вы математики, если запаролиться нормально не можете. 8426 —  | 7329 —
| 7329 —  или читать все.
или читать все.
Чтобы перейти к разговору о радиальном транспорте воды, необходимо коротко вспомнить общие черты строения корней (хотя в деталях строение корней в разных видах растений варьирует). Упрощенная общая схема поперечного разреза корня однодольных и двудольных растений представлена на рис 4.10.

Рис. 4.10. Схематическое изображение поперечного разреза корня
В обоих типах растений имеется центральный цилиндр (стела), в котором находятся сосудистые пучки и (стелярная) паренхима. При помощи сосудистых пучков (флоэма) поддерживается связь между разными частями растения и местами синтеза углеводов. По сосудистым пучкам (ксилема) поступает из корня в побег вода и растворенные вещества. Если перемещаться к периферии, то далее идет перецикл – однорядный слой клеток. Затем идет эндодерма – тоже однорядный слой клеток, особенно хорошо приспособленный к тому, чтобы служить барьером, который отделяет проводящие ткани от коры. После того, как растяжение клеток заканчивается, в радиальных и поперечных оболочках эндодермы выявляется прослойка материала, которая дает положительную реакцию на лигнин. Это так называемый поясок Каспари. Пояски Каспари сильно ограничивают движение воды, ионов в клеточных оболочках. Таким образом, они эффективно блокируют апопластический транспорт.
Большую часть корней занимают клетки коры. На их долю (вместе с эпидермисом) приходится 86–90 % площади. Клетки коры вытянуты параллельно главной оси, тонкий слой цитоплазмы (1–5 мкм) окружает в них центральную вакуоль, которая занимает
90 % объема протопласта. Вторичные изменения в оболочках клетки коры сводятся к отложению целлюлозы; только у некоторых видов происходит одревеснение. Вообще, оболочки сохраняют проницаемость. В них имеются многочисленные поры, которые могут быть заполнены в зависимости от условий воздухом или водой.
По периферии корня расположен эпидермис (ризодермис). Это наружная ткань корня, состоящая из вытянутых плотно упакованных клеток. Оболочки этих клеток могут со временем претерпевать изменения, связанные с отложением кутина – водоотталкивающего вещества.
Внешние тангенциальные оболочки клеток ризодермиса способны сильно растягиваться и образовывать трубковидные выросты, так называемые корневые волоски. Отметим, что клетки, которые могут образовывать корневые волоски имеют название трихобластов, а которые не способны к образованию – атрихобластов.
Итак, вода, попавшая в корневой волосок или другую клетку корня с помощью одного из механизмов, о которых мы говорили, движется к сосудам ксилемы, т. е. происходит радиальное перемещение воды по тканям корня (рис. 4.11).
Движение воды в корневой волосок или другую клетку начинается с взаимодействия с оболочкой клеток, затем вода может пройти через плазмалемму и транспортироваться из протопласта одной клетки в протопласт другой через плазмодесмы. В этом случае вода движется по симпласту. Однако вода могла бы и не заходить в симпласт, а оставаться в клеточной оболочке и двигаться по ним через ткани корня к центральному цилиндру.

Рис. 4.11.Путь воды от корневого волоска (1) до сосудов (12) через клетки коры (2 – 6), эндодерму (7), перицикл (8) и паренхиму осевого цилиндра (9 – 11)
Это апопластический путь. Но по апопласту вода может двигаться только до эндодермы (до поясков Каспари). Поэтому путь воды по апопласту тут прекращается. Для дальнейшего передвижения она должна обязательно войти в симпласт.
Первым барьерную функцию эндодермы выявил Д. Пристли в 1920 г. В последнее время эта функция была неоднократно подтверждена.
Необходимость переключения движения воды с апопластного на симпластный путь имеет большое значение, так как процесс движения по живой протоплазме клетка может сама регулировать в отличие от перемещения воды по клеточным стенкам.
Для поступления воды в корень необходимо чтобы водный потенциал ризодермиса был меньше, чем водный потенциал почвенного раствора; в этом случае вода начнет поступать в клетки.
Почву можно рассматривать как резервуар, количество воды, в котором то увеличивается, то уменьшается. Если после дождя почва находится в состоянии полевой влагоемкости, то ее водный потенциал близок к нулю; вода легко поступает в корни. По мере подсыхания почвы ее водный потенциал снижается.
Водный потенциал максимален в почве, несколько меньший в клетках корня и самый низкий в клетках, прилегающих к эпидермису листа (табл. 4.2).
Таблица 4.2. Величины водного потенциала в системе почва-растение-воздух
Водный потенциал, бар
Первая причина возникновения градиента водного потенциала – активное поступление солей и их активное перемещение из одной клетки в другую. Вторая – живые клетки паренхимы центрального цилиндра выделяют растворимые органические и минеральные вещества в сосуды и таким образом поддерживают градиент водного потенциала в сосудах. Поступая в сосуды ксилемы, они образуют осмотический градиент.
Химический потенциал воды, в которой находятся эти ионы ниже потенциала воды в разбавленном солевом растворе внешней среды. Поэтому вода должна перемещаться по градиенту водного потенциала в ксилему корня.
Сегодня, выдвинута гипотеза, которая имеет под собой экспериментальную основу, что в корнях есть специализированный механизм активного накачивания воды (водная помпа), работа которой не зависит от поступления ионов.
Действительно, для поступления воды в корень необходима энергия, поэтому этот процесс зависит от скорости аэробного дыхания корней – основного источника АТФ. Таким образом, одним из главных факторов, обуславливающих поступление воды, является О2. Отсюда условия, способные подавить дыхание, резко уменьшают поступление воды. В качестве примера рассмотрим следующее явление: прошел сильный дождь, в низком месте собралось много воды, а растение завядает. Почему? Избыток воды в почве вытеснил из нее воздух, поступление кислорода в корень затруднено, дыхание подавлено. Сразу же наблюдается торможение поступления воды в корневую систему. Этот пример объясняет, почему на затопленных водой почвах растения развиваются плохо и даже гибнут.
Затопление приводит не только к уменьшению количества О2, но и к увеличению концентрации СО2 в почве, который повреждает мембраны корневых волосков; наблюдается снижение поступления воды, что подтверждается торможением выделения пасоки.
Поступление воды в корень зависит и от температуры почвы. Например, на холодных болотистых почвах, несмотря на большое количество воды, растения испытывают недостаток воды, поскольку при низких температурах подавляется дыхание и нарушается снабжение корней энергией. У растений в этих условиях начинает формироваться ксероморфная структура (мелкие клетки, много устьиц и т. д.), которая характерна для сухих мест.
Состояние растений, при котором вода не может поступать, несмотря на ее большое количество в окружающей среде, получило название физиологической засухи.
Различные полютанты также могут влиять на поступление воды, в частности через подавление дыхания.
Необходимость поглощать большое количество воды даже в условиях ее лимита, например, в условиях засухи, приводит к тому, что растение формирует огромную корневую систему. В результате корни проникают в почву на большую глубину. У пшеницы длина корней достигает 90 см, у люцерны – 120 см. Если считать, что глубина пахотного горизонта составляет 20–25 см, то большая часть корневой системы размещается ниже. Размер корневой системы характеризуется не только глубиной их проникновения в почву. Большое значение имеет общая поверхность.
Вода в почве двигается очень медленно: на протяжении месяца она продиффундирует не больше, чем на 30 см. Перемещение кончика корня в почве опережает движение воды. Таким образом, не вода движется к корню, а корень к воде в процессе роста. Рост является первой и важнейшей особенностью корней как органа, поглощающего воду.
В засушливых условиях формируется корневая система в 3–4 раза большая, чем во влажных.
Разветвление и быстрый рост помогают корню двигаться к воде, но, с другой стороны, вода – необходимое условие роста. Как видно уже не на клеточном, а на организменном уровне, мы сталкиваемся с примером обратной связи, которая лежит в основе регуляторных процессов.
Как орган, поглощающий воду, корень владеет еще одним важным свойством – положительным гидротропизмом, это значит, что при недостатке воды растущие части корней выгибаются в стороны более влажных участков почвы.
Таким образом, корневая система представляет собой специализированный орган поступления воды.
Рост корней обычно опережает рост наземных органов. Это очень важная особенность, связанная с тем, что корень должен обеспечить потребности в воде формирующего растения.
Однако клетка любого органа, которая не насыщена водой, тоже может поглощать воду, как только она будет приведена с ней в соприкосновение. Поэтому и листья, особенно подвядшие, при погружении в воду довольно энергично ее всасывают; с другой стороны, несмотря на кутикулу, вода может поступать через поверхность листа. Показано, что только сухая кутикула почти непроницаема для воды; при смачивании она набухает и делается проницаемой, поэтому смоченные дождем или росой листья могут поглощать до 25 % падающей на них воды. Это имеет практическое значение при орошении растений дождеванием.
На рисунке показано первичное строение корня типичного двудольного растения.
Основная масса воды поглощается более молодыми частями корня в зоне корневых волосков. По мере роста корня в почве на нем постоянно образуются новые корневые волоски, а старые отмирают. Новые корневые волоски возникают на некотором расстоянии позади зоны растяжения. Эти волоски представляют собой трубчатые выросты эпидермальных клеток (рис. 13.17), существенно увеличивающие поверхность, способную поглощать воду и минеральные соли. Они вступают в тесный контакт с частицами почвы.
На рисунке схематично показаны пути движения воды по корню. В корне существует градиент водного потенциала — от более высокого в клетках, образующих корневые волоски, к более низкому в клетках, примыкающих к ксилеме. Этот градиент поддерживается двумя способами:
1) за счет движения воды вверх по ксилеме, при котором, как мы уже говорили, в ксилеме создается натяжение (отрицательное давление) и тем самым понижается водный потенциал ксилемного сока;
2) за счет того, что осмотический потенциал ксилемного сока более низкий (более отрицательный) по сравнению с осмотическим потенциалом разбавленного почвенного раствора.
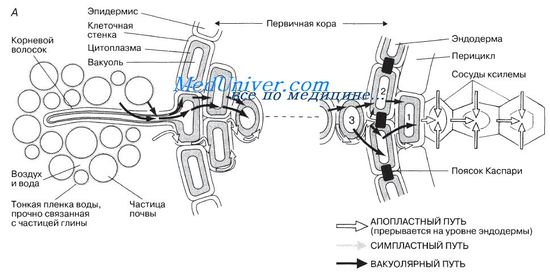
Вода движется через корень по тем же путям, что и в листьях, а именно по апопласту, симпласту и через вакуоли.
По мере того как вода поднимается вверх по корневой ксилеме, ее замещает вода из окружающих паренхимных клеток, например из клетки 1 на рисунке. В результате водный потенциал этой клетки снижается и в нее устремляется вода из соседней клетки 2 благодаря осмосу или просто по симпласту, как описано в разд. 13.3.2 для мезофилла листа. Тогда в свою очередь снижается и водный потенциал клетки 2, в нее начинает поступать вода из клетки 3 и так далее через весь корень до самого эпидермиса, образующего волоски.
Водный потенциал почвенного раствора выше, чем в клетках эпидермиса и в корневых волосках. Следовательно, вода проникает в корень извне путем осмоса.
Апопластный транспорт в корне
Апопластный транспорт в корне происходит примерно так же, как в листьях, но с одним существенным отличием. Когда вода, продвигаясь по клеточным стенкам, достигает эндодермы, путь ей преграждает водонепроницаемое вещество, называемое суберином. Оно откладывается по периметру эндодермальной клетки в ее антиклинальных стенках, образуя так называемый поясок Каспари (рис. 13.18, Б). В результате вода с растворенными в ней веществами (в основном диссоциированными на ионы солями) должна сначала проникнуть через плазмалемму этой клетки в ее цитоплазму, а потом выйти «с другой стороны». Таким способом клетки эндодермы контролируют и регулируют движение растворов по пути к ксилеме. Такой контроль необходим для защиты побегов от проникновения в них токсичных веществ, бо-лезнетвореных бактерий, грибов и других вредных агентов. С возрастом отложение суберина в эндодермальных клетках корня увеличивается, и это препятствует нормальному выходу воды и растворенных солей через внутренние танген-тальные клеточные стенки (рис. 13.18, Б). Однако в таких стенках могут сохраняться поры и проходящие через них плазмодесмы и, кроме того, остаются так называемые «пропускные» клетки, у которых не происходит дополнительного утолщения стенки и через которые свободно проходят вода и растворенные вещества. Количественное соотношение в корне апопластного, симпластного и вакуолярного транспортов воды не известно.
